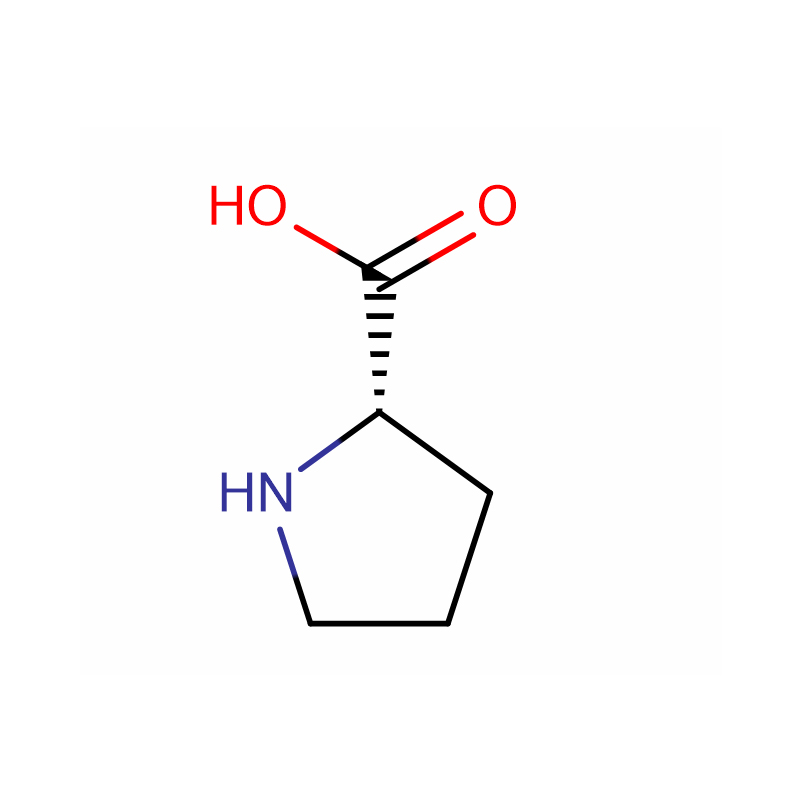
L-Proline Cas: 147-85-3 99% Polvo blanco
| Numero de catalogo | XD90293 |
| nombre del producto | L-prolina |
| CAS | 147-85-3 |
| Fórmula molecular | C5H9NO2 |
| Peso molecular | 115.13046 |
| Detalles de almacenamiento | Ambiente |
| Código Arancelario Armonizado | 29339980 |
Especificaciones del producto
| Ensayo | 99%mín. |
| Apariencia | polvo blanco |
| Rotacion especifica | -84,5 a -86 |
| Metales pesados | <15 ppm |
| AS | <1ppm |
| Ph | 5,9 - 6,9 |
| SO4 | <0.050% |
| Fe | <30 ppm |
| Pérdida por secado | <0.3% |
| Residuos en ignición | <0.10% |
| NH4 | <0.02% |
| Cl | <0.050% |
| Estado de solución | >98% |
Comprender el metabolismo del huésped microbiano es esencial para el desarrollo y la optimización de los procesos biocatalíticos basados en células completas, ya que determina la eficiencia de la producción.Esto es especialmente cierto para la biocatálisis redox donde se emplean células metabólicamente activas debido a la capacidad regenerativa del cofactor/cosustrato endógeno en el huésped.Se usó Escherichia coli recombinante para sobreproducir prolina-4-hidroxilasa (P4H), una dioxigenasa que cataliza la hidroxilación de L-prolina libre en trans-4-hidroxi-L-prolina con a-cetoglutarato (a-KG) como cosustrato.En este biocatalizador de células enteras, el metabolismo central del carbono proporciona el cosustrato requerido a-KG, acoplando el rendimiento biocatalítico de P4H directamente al metabolismo del carbono y la actividad metabólica.Mediante la aplicación de herramientas de biología experimental y computacional, como la ingeniería metabólica y el análisis de flujo metabólico de (13) C ((13) C-MFA), investigamos y describimos cuantitativamente la respuesta fisiológica, metabólica y bioenergética del biocatalizador de células enteras. a la bioconversión dirigida e identificó posibles cuellos de botella metabólicos para una mayor ingeniería de vías racionales. Se construyó una cepa de E. coli deficiente en la degradación de prolina eliminando el gen putA que codifica la prolina deshidrogenasa.Las biotransformaciones de células completas con esta cepa mutante condujeron no solo a la hidroxilación cuantitativa de prolina, sino también a una duplicación de la tasa de formación específica de trans-4-L-hidroxiprolina (hyp), en comparación con el tipo salvaje.El análisis del flujo de carbono a través del metabolismo central de la cepa mutante reveló que el aumento de la demanda de a-KG para la actividad de P4H no mejoró el flujo de generación de a-KG, lo que indica una operación del ciclo TCA estrechamente regulada en las condiciones estudiadas.En la cepa de tipo salvaje, la síntesis y catálisis de P4H provocaron una reducción en el rendimiento de biomasa.Curiosamente, la cepa ΔputA compensó adicionalmente la pérdida asociada de ATP y NADH al reducir las demandas de energía de mantenimiento a tasas de captación de glucosa comparativamente bajas, en lugar de aumentar la actividad de TCA. ser prometedor para la catálisis productiva de P4H no solo en términos de rendimiento de biotransformación, sino también con respecto a las tasas de biotransformación y absorción de prolina y el rendimiento de hip en la fuente de energía.Los resultados indican que, tras una inactivación de putA, el acoplamiento del ciclo TCA a la hidroxilación de prolina a través del cosustrato a-KG se convierte en un factor limitante clave y un objetivo para mejorar aún más la eficiencia de las biotransformaciones dependientes de a-KG.